La luz viaja por los objetos transparentes a menor velocidad que el aire. El índice de refracción de una sustancia transparente es el cociente entre la velocidad de la luz en el aire y la velocidad en ese objeto transparente.

El sentido de la luz es siempre perpendicular al plano del frente de onda. Cuando una onda luminosa atraviesa una superficie angulada, se refracta (dobla) con un cierto ángulo si los índices de refracción de los dos medios son diferentes. El ángulo depende del índice de refracción del material de barrera y del ángulo entre las dos superficies.
Aplicación de los principios de la refracción a las lentes.
Una lente convexa concentra los rayos de luz. Los rayos de luz que atraviesan el perímetro de la lente se doblan (refractan) hacia los que atraviesan la región central (hasta volverse perpendiculares al frente de onda). Se dice que los rayos de luz convergen.
Una lente cóncava dispersa los rayos de luz. En el perímetro de la lente, las ondas luminosas se refractan, de manera que viajan perpendiculares al frente de onda, o interfaz, y se doblan, desviándose de las que atraviesan la región central. Este fenómeno se llama divergencia.
La distancia focal de una lente es la situada más allá de una lente convexa, en la que los rayos de luz paralelos convergen en un punto común. Cada foco puntual de luz situado delante de una lente convexa se concentra al otro lado de la lente, alineado con su centro, es decir, el objeto se ve boca abajo e invertido de izquierda a derecha. Cuanto más se doblan los rayos de luz por una lente, mayores su poder de refracción. La unidad de medida del poder de refracción es la dioptría. Una lente esférica (o convexa) que convergiera los rayos de luz paralelos en un punto situado1 metro más allá de la lente posee un poder de refracción de+1 dioptría, y si los rayos se refractan dos veces más, se dice que la potencia es de +2 dioptrías.
Óptica del ojo
El ojo, desde el punto de vista óptico, equivale a una cámara fotográfica. Posee una lente, una apertura variable (la pupila) y la retina, que se corresponde con la película. El sistema de lentes del ojo enfoca una imagen invertida (boca abajo) sobre la retina. Sin embargo, la imagen se percibe correctamente, porque el cerebro ha «aprendido» que esa es la orientación correcta o normal. La acomodación depende de un cambio en la forma del cristalino y permite que el ojo enfoque los objetos cercanos. Cuando se desvía la mirada de un objeto lejano a otro próximo, aparece el proceso de acomodación, que:1) aumenta la convexidad del cristalino; 2) estrecha el diámetro de la pupila, y 3) produce una aducción (vergencia) de los dos ojos. Si el cristalino se encuentra en un estado «relajado», sin que se ejerza ninguna tensión en los bordes de su cápsula, adopta una forma casi esférica en virtud de sus propiedades elásticas intrínsecas. Cuando las fibras inelásticas de la zónula, adheridas al perímetro del cristalino, se tensan y traccionan radialmente desde su inserción en el músculo ciliar inactivo (y en el cuerpo ciliar), el cristalino se vuelve relativamente plano o menos convexo. Si se activa el músculo ciliar por las fibras parasimpáticas posganglionares del nervio motor ocular comunes, se contraen las fibras circulares del músculo ciliar, que producen una acción de tipo esfinteriano relajando la tensión sobre las fibras zonulares y permitiendo una mayor convexidad del cristalino como consecuencia de su elasticidad inherente. Con ello aumenta la capacidad de refracción y el ojo puede enfocar los objetos cercanos. Al mismo tiempo, el músculo esfínter de la pupila se activa, se contrae la pupila y se produce una desviación interna de ambos ojos.
La presbicia es la pérdida de acomodación por el cristalino. Cuando se envejece, el cristalino empieza a perder sus propiedades elásticas intrínsecas y responde peor, volviéndose incapaz de enfocar los objetos cercanos. Este trastorno (presbicia) se corrige con gafas de lectura, diseñadas para aumentar los objetos cercanos, o con lentes bifocales, en las que la parte superior aumenta la visión lejana, y la inferior, la capacidad de refracción, a fin de mejorar la visión de cerca.

El diámetro de la pupila (iris) también contribuye a la acomodación. Cuanto mayor es el diámetro, más luz penetra en el ojo. Al entrecerrar los ojos (estrechar el orificio pupilar), mejora la nitidez de la imagen, porque aumenta el plano focal.
Errores de refracción
La emetropía se refiere al estado normal del ojo. Cuando el músculo ciliar se encuentra completamente relajado, todos los objetos alejados se enfocan con nitidez en la retina.
La hipermetropía se debe a que el globo ocupar es demasiado corto desde su vértice hasta la base, por lo que los rayos de luz son enfocados detrás de la retina; este trastorno se corrige con una lente convexa.

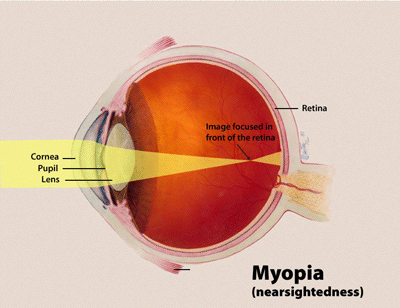
La miopía se debe a que el globo ocular se alarga de delante hacia atrás y, por eso, los rayos de luz se enfocan delante de la retina; este trastorno se corrige con una lente cóncava, que disminuye la refracción y ocasiona una divergencia de los rayos de luz incidentes.
El astigmatismo se debe a diferencias considerables en la curvatura de los diferentes planos oculares. Así, la curvatura en el plano vertical del ojo puede ser bastante menor que la horizontal. El resultado es que los rayos de luz que inciden en el ojo desde direcciones diferentes se enfocan en puntos diferentes. Para corregir este trastorno se necesita una lente cilíndrica. Las cataratas se deben a la opacidad que se forma en una parte del cristalino. El tratamiento preferido es la extracción del cristalino y su sustitución por una lente artificial. El queratocono es un trastorno causado por una forma extraña de la córnea, que sobresale mucho a un lado y provoca un problema de refracción grave que no puede corregirse con una sola lente. La mejor solución pasa por una lente de contacto, adherida a la superficie corneal y mantenida en el sitio por una película del líquido lagrimal. La lente se esmerila para compensar la protrusión de la córnea, de manera que la cara anterior de la lente de con-tacto se transforma en una superficie de refracción mucho más uniforme y eficaz.
Agudeza visual: más nítida en la fóvea de la retina
La fóvea se compone enteramente de conos fotorreceptores, cada uno con un diámetro aproximado de 1,5 mm. La agudeza visual normal de las personas facilita la discriminación entre dos puntos de luz si estos se encuentran separados en la retina por un arco mínimo de 25 s. El diámetro normal de la fóvea es de unos 0,5 mm. La agudeza máxima se concentra en un campo visual menor de2 grados. El descenso de la agudeza fuera de la fóvea se debe, en parte, a la presencia de bastones mezclados con conos y a la unión de algunos bastones y conos a las mismas células ganglionares. Los optotipos para explorar la agudeza visual suelen colocarse a 6 m de la persona examinada. Si se reconocen las letras de un determinado tamaño a 6 m, se dice que la visión de esa persona es de 20/20. Si, a 6 m, la persona solo ve letras que deberían verse a 60 m, se dice que tiene una visión de 20/200.
Determinación de la distancia de un objeto al ojo: «percepción de la profundidad»
Si se conoce el tamaño de un objeto, el cerebro puede calcular su distancia con respecto al ojo. Si una persona mira a un objeto remoto sin mover los ojos, no ocurre ningún movimiento de paralaje. Sin embargo, si la cabeza se des plaza de un lado a otro, los objetos próximos se mueven rápidamente a través de la retina, mientras que los lejanos lo hacen muy poco o nada. La visión binocular también ayuda a determinar la distancia de un objeto. Como los ojos suelen estar separados unos5 cm, un objeto situado a 2,5 cm del puente de la nariz sería visto por una pequeña parte de la retina periférica de ambos ojos. En cambio, la imagen de un objeto situado a 6 m incide en puntos prácticamente correspondientes del centro de cada retina. Este tipo de paralaje binocular (estereopsia) permite establecer con precisión las distancias con relación al ojo.
El oftalmoscopio
El oftalmoscopio ilumina la retina del ojo observado por medio de un espejo o prisma angulado y una pequeña bombilla. El observador coloca el instrumento para ver la retina del sujeto a través de la pupila. Si los ojos del sujeto o los del examinador no son emétropes, la refracción se puede ajustar con una serie de lentes movibles del oftalmoscopio.
Sistema humoral del ojo: líquido intraocular
El humor vítreo se encuentra entre el cristalino y la retina yes un cuerpo más gelatinoso que líquido. Las sustancias pueden difundir a su través, pero el movimiento o flujo de este líquido es mínimo. El humor acuoso es un líquido acuoso segregado por el epitelio de las prolongaciones ciliares del cuerpo ciliar con una velocidad de 2 a 3 ml/min. Este líquido emigra entre los ligamentos de sujeción del cristalino y atraviesa la pupila para alcanzar la cámara anterior del ojo (situada entre el cristalino y la córnea). Desde aquí, el líquido fluye hasta el ángulo formado por la córnea y el iris, y luego atraviesa una malla trabecular, entrando en el conducto de Schlemm, que desemboca directamente en las venas extraoculares. La presión intraocular normal es de unos 15 mmHg, y varía entre 12 y 20 mmHg. De ordinario, se emplea un tonómetro para medir la presión intraocular. Este aparato se compone de una pequeña pletina que se coloca sobre la córnea anestesiada. Se aplica una ligera fuerza sobre la pletina, que desplaza la córnea hacia dentro, y se calibra la distancia interna recorrida en forma de presión intraocular. El glaucoma es un trastorno en el que la presión intraocular puede alcanzar cifras peligrosamente elevadas (del orden de 60 a 70 mmHg). A medida que la presión asciende por encima de 20 a 30 mmHg, los axones de las células ganglionares de la retina que forman el nervio óptico se comprimen hasta el extremo de interrumpir el flujo axónico, causando un daño permanente de la neurona originaria. La compresión dela arteria central de la retina también puede producir una muerte neuronal en la retina. El glaucoma se puede tratar con colirios oculares, que reducen la secreción del humor acuoso o que aumentan su absorción. Si fracasa la farmacoterapia, se aplican intervenciones quirúrgicas para abrir los espacios trabeculares o para drenar directamente la malla trabecular hacia los espacios subconjuntivales situados fuera del globo ocular.
El ojo: II. Función receptora y nerviosa de la retina
Anatomía y función de los elementos estructurales de la retina
La retina se compone de 10 capas o estratos celulares que se enumeran de forma secuencial, comenzando por el más externo (el más alejado del centro del globo ocular):
1. Capa pigmentaria
2. Capa de conos y bastones
3. Membrana limitante externa
4. Capa nuclear externa
5. Capa plexiforme externa
6. Capa nuclear interna
7. Capa plexiforme interna
8. Capa ganglionar
9. Capa de fibras del nervio óptico
10. Membrana limitante interna

Cuando la luz atraviesa el sistema ocular de lentes, encuentra primero la membrana limitante interna, las fibras del nervio óptico y la capa de células ganglionares; luego prosigue por las demás capas hasta alcanzar los receptores (conos y bastones). La fóvea es una región especializada, de aproximadamente 1 mm2, situada en el centro de la retina. Dentro del centro de la fóvea existe una zona, denominada fóvea central, con un diámetro de 0,3 mm; esta es la región donde la agudeza visual alcanza el máximo y la capa de fotorreceptores solo contiene conos. Además, las capas subyacentes de la retina a lo largo de las fibras del nervio óptico y de los vasos sanguíneos– se desplazan lateralmente para facilitar el acceso más directo posible a los receptores. Cada fotorreceptor se compone de: 1) un segmento externo; 2) un segmento interno; 3) una región nuclear, y 4) el cuerpo o terminal sináptico. Los receptores se denominan bastones o conos, dependiendo, principalmente, de la forma del segmento externo. El fotopigmento sensible a la luz rodopsina se localiza en el segmento externo del bastón, mientras que el del cono con-tiene un material parecido, un pigmento sensible a los colores, la fotopsina. Estos fotopigmentos son proteínas incorporadas aun conjunto apilado de discos membranosos en el segmento externo del receptor, que representa un repliegue de la membrana celular externa. Sin embargo, no resulta fácil de reconocer en la porción distal del segmento externo del bastón, donde los discos membranosos se desprenden de manera secundaria y están totalmente contenidos dentro de la membrana limitante de ese segmento. Los segmentos internos de los bastones y conos son básicamente idénticos y contienen componentes y orgánulos citoplásmicos comunes a otros somas neuronales. Cada núcleo del fotorreceptor se continúa con su propio segmento interno, pero la membrana limitante externa de la retina establece una separación o frontera incompleta entre la capa de los segmentos internos y la capa de los núcleos fotorreceptores (capa nuclear externa).El cuerpo sináptico contiene elementos como mitocondrias y vesículas sinápticas, habituales en las terminaciones axónicas del cerebro. El pigmento negro melanina de la capa de pigmento reduce la reflexión de la luz por el globo ocular y cumple, por tanto, una función parecida al color negro del interior de los fuelles de una cámara. La importancia de este pigmento se reconoce mejor por su ausencia en las personas albinas. Los albinos casi nunca alcanzan una agudeza visual mayor de 20/100 debido a la enorme reflexión que tiene lugar dentro del globo ocular. La capa de pigmento también alma-cena grandes cantidades de vitamina A, empleadas en la síntesis de los pigmentos visuales. La arteria central de la retina irriga solo las capas más internas (desde los axones de las células ganglionares hasta la capa nuclear interna). Las capas más externas de la retina son perfundidas mediante difusión desde las coroides, tejido muy vascularizado que se sitúa entre la esclerótica y la retina. Cuando una persona sufre un desprendimiento traumático de retina, la línea de separación ocurre entre la retina nerviosa y el epitelio pigmentario. Dada la perfusión independiente de las capas internas de la retina a través de la arteria central, la retina sobrevive varios días y puede resistirse a la degeneración funcional si se consigue su aposición normal al epitelio pigmentario en el quirófano.

Fotoquímica de la visión
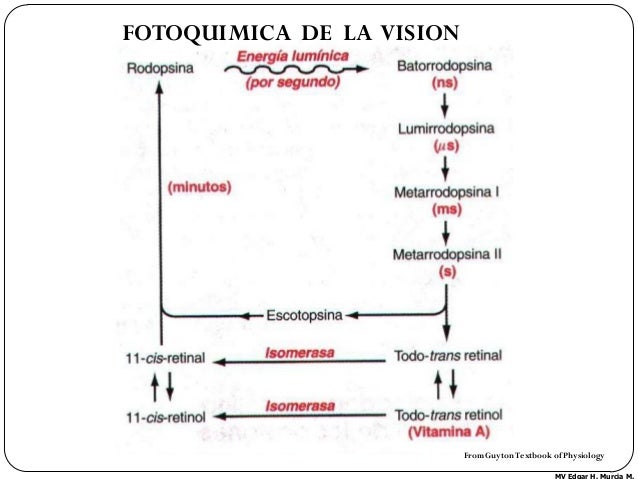
Ciclo rodopsina retinal y excitación de los bastones. La rodopsina se descompone por la energía lumínica. El fotopigmento rodopsina de los bastones se concentra en la porción del segmento externo que sobresale hacia la capa pigmentaria. Esta sustancia es una combinación de la proteína escotopsina y del pigmento carotenoide retinal o, más concretamente,11-cis retinal. Cuando la rodopsina absorbe la energía lumínica, el retinal se transforma en su configuración todo- trans y comienzan a separarse los componentes retinal y escotopsina. Tras una serie de reacciones extraordinariamente rápidas, el componente retinal se transforma en lumirrodopsina, metarrodopsina I, metarrodopsina II y, por último, escotopsina; y se escinde el todo-trans retinal. Durante este proceso, se cree que la metarrodopsina II se ocupa de la transmisión de los impulsos sucesivos por la retina. Regeneración de la rodopsina. En la primera etapa de regeneración de la rodopsina, el todo-trans retinal se con-vierte en 11-cis retinal, que se combina en seguida con la escotopsina para dar la rodopsina. Existe una segunda vía deformación de la rodopsina basada en la conversión del todo-trans retinal en todo-trans retinol, una forma de vitamina A. El retinol se convierte por vía enzimática en 11-cis retinol y luego en 11-cis retinal, que puede combinarse con la escotopsina para dar rodopsina. Si la retina contiene un exceso de retinal, se transforma en vitamina A, disminuyendo así la cantidad total de rodopsina de la retina. Las personas con carencia de vitamina A experimentan ceguera nocturna porque los bastones son los fotorreceptores que más se utilizan en condiciones de iluminación crepuscular y la formación de rodopsina disminuye de forma espectacular por la ausencia de vitamina A. Este trastorno se revierte en 1 h o menos mediante la inyección intravenosa de vitamina A.
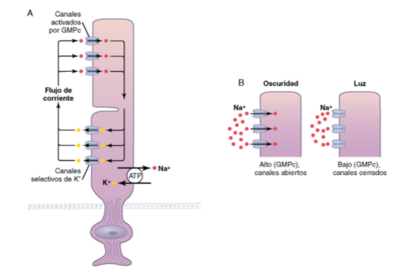
Excitación del bastón cuando la luz activa la rodopsina
Los bastones siguen un comportamiento muy distinto a otros receptores neurales. En la oscuridad (sin ninguna estimulación lumínica), las membranas del segmento externo de los bastones son «permeables» al sodio, es decir, los iones sodio entran en el segmento externo y cambian el potencial de membrana desde el valor característico de –70 a –80 mV, hallado en los receptores sensitivos, hasta un valor más positivo de –40 mV. Esta se denomina corriente de sodio o «corriente oscura» y hace que se libere una pequeña cantidad de transmisor en la oscuridad. Cuando la luz incide en el segmento externo del bastón, las moléculas de rodopsina experimentan la serie de reacciones expuesta con anterioridad, lo que reduce la conductancia del sodio por el segmento externo y también la corriente oscura. Algunos iones sodio siguen siendo bombeados fuera de la membrana celular y la pérdida de estos iones positivos hace que el interior de la célula se torne más negativo; el potencial de membrana se hace más negativo, es decir, se hiperpolariza. Se detiene entonces el flujo del transmisor. Cuando la luz incide sobre un fotorreceptor, la hiperpolarización pasajera de los bastones alcanza el máximo en unos 0,3 s y dura más de 1 s. Además, la magnitud del potencial de receptor es proporcional al logaritmo de la intensidad lumínica. Este hecho reviste enorme significado funcional, porque permite que el ojo discrimine la intensidad lumínica en un intervalo muchos miles de veces superior al teórico. Todo ello es consecuencia de una cascada química extremadamente sensible que multiplica casi un millón de veces los efectos estimuladores de la siguiente manera. La rodopsina activada (metarrodopsina II) actúa, a modo de enzima, activando numerosas moléculas de transducina, una proteína que se encuentra también en la membrana de los discos del segmento externo. La transducina activada activa, a su vez, la fosfodiesterasa, enzima que hidroliza de inmediato múltiples moléculas de monofosfato de guanosina cíclico (GMPc). Esta pérdida de GMPc hace que se cierren muchos canales de sodio, con lo que el potencial de membrana se va volviendo cada vez más negativo (hiperpolarizado). En el plazo de 1 s, la metarrodopsina II se inactiva y toda la cascada revierte: el potencial de membrana se torna más despolarizado a medida que se vuelven a abrir los canales de sodio, y el sodio, una vez más, penetra en el segmento externo al restablecerse la corriente oscura. Los conos se comportan de forma parecida, pero el factor de amplificación es entre 30 y 300 veces menor que el de los bastones.
Fotoquímica de la visión de los colores por los conos
Como sucede con los bastones ya descritos, la transducción fotoquímica en los conos depende de una opsina y de un retinal. La opsina de los conos se denomina fotopsina, y su composición química es distinta a la de la rodopsina, mientras que el componente retinal es exactamente el mismo que el delos bastones. Existen tres tipos de conos, caracterizados cada uno por una fotopsina diferente con una sensibilidad máxima a una longitud de onda determinada de la luz, en la porción azul, verde o roja del espectro lumínico.

Adaptación a la luz y a la oscuridad
Si se efectúa una exposición prolongada a una luz brillante, un porcentaje considerable de las sustancias fotoquímicas de los bastones y de los conos disminuye y gran parte del retinal se transforma en vitamina A. El resultado es un des-censo de la sensibilidad global a la luz, que se conoce como adaptación a la luz. Por el contrario, si una persona permanece mucho tiempo en la oscuridad, las opsinas y el retinal se convierten de nuevo en pigmentos fotosensibles. Además, la vitamina A se transforma en retinal, aportando aún más pigmento fotosensible; este proceso se denomina adapta-ión a la oscuridad y tiene lugar cuatro veces más de prisa en los conos que en los bastones, pese a que los primeros resulten menos sensibles a la oscuridad. Los conos cesan de adaptarse a los pocos minutos, mientras que los bastones, con una adaptación más lenta, continúan haciéndolo durante minutos u horas, y su sensibilidad aumenta dentro de un amplio espectro. La adaptación puede ocurrir, asimismo, a través de cambios del tamaño pupilar, que puede variar del orden de 30 veces en una fracción de segundo. Los circuitos existentes dentro dela retina y del cerebro también presentan una adaptación neural. Cuando aumenta la intensidad lumínica, también lo hace la transmisión desde la célula bipolar hasta la célula horizontal, la célula amacrina y la célula ganglionar. A pesar de que esta última adaptación es menos potente que los cambios pupilares, la adaptación neural, como la pupilar, ocurre con rapidez. La utilidad de la adaptación a la luz y a la oscuridad permite que el ojo experimente cambios de sensibilidad por factores de hasta 500.000 a 1 millón, como se reconoce cuando, desde un ambiente muy iluminado, se pasa a una sala oscura. La sensibilidad de la retina es baja, porque está adaptada a la luz y en una habitación oscura se ve poco. Cuando se produce la adaptación a la oscuridad, mejora la visión en esas condiciones. Se cree que la intensidad de la luz solar es 10.000 millones de veces mayor que la intensidad de la luz en una noche estrellada. Sin embargo, el ojo puede operar, hasta cierto punto, en ambas condiciones, debido a su enorme margen de adaptación.


Visión en color
Mecanismo tricolor para la detección del color
La sensibilidad espectral de los tres tipos de conos depende delas curvas de absorción de la luz de los tres pigmentos respectivos. Todo el color visible (aparte del azul, verde o rojo) es el resultado de la estimulación combinada de dos o más tipos de conos. El sistema nervioso interpreta como color la relación entre la actividad de los tres tipos. Se interpreta como luz blanca una estimulación equivalente de los conos azules, verdes y rojos. Si se modifica el color de la luz que ilumina una escena, no se altera sustancialmente la tonalidad del color de esa escena, lo que se conoce como constancia del color, mecanismo atribuido a la corteza visual primaria. Cuando falta un tipo concreto de cono de la retina, no se logran distinguir ciertos colores. Una persona sin conos rojos sufre protanopía. El espectro global se acorta por el extremo largo de la longitud de onda cuando faltan los conos rojos. La ceguera para los colores rojo y verde es un defecto genético de los hombres transmitido por las mujeres. Los genes del cromosoma X femenino codifican los conos respectivos. Este defecto es raro en las mujeres, porque posee dos cromosomas X y casi siempre disponen de una copia normal del gen.

Función nerviosa de la retina
Circuitos nerviosos de la retina
Los fotorreceptores se componen de los segmentos externos de los bastones y conos y de los segmentos internos de la capa de fotorreceptores, el cuerpo celular de la capa nuclear externa, y el cuerpo sináptico de la capa plexiforme externa. Las células horizontales, las células bipolares y las células amacrinas reciben aferencias sinápticas de la capa plexiforme externa, poseen somas en la capa nuclear interna y establecen contactos presinápticos con la capa plexiforme interna. Las células ganglionares reciben aferencias sinápticas de la capa plexiforme interna, posee en el soma en la capa de células ganglionares y originan axones que viajan dentro de la capa de fibras del nervio óptico. Las células interplexiformes transmiten las señales en sentido contrario, desde la capa plexiforme interna hasta la externa. En la fóvea, la ruta desde el cono hasta la célula ganglionares bastante directa y puede consistir en un receptor, una célula bipolar y una célula ganglionar. Las células horizontales pueden intervenir en la capa plexiforme externa, pero las amacrinas ejercen su actividad en la capa plexiforme interna. En las zonas más periféricas de la retina, donde abundan más los bastones, las señales de varios fotorreceptores pueden converger sobre una única neurona bipolar, cuya salida podría dirigirse únicamente a una célula amacrina que se proyecta después hasta una célula ganglionar. Esta constituye la vía dela visión pura con los bastones. Las células horizontales y amacrinas proporcionan una conectividad lateral. Los neurotransmisores presentes en la retina son el glutamato (utilizado por los bastones y los conos) y el ácido gaminobutírico (GABA), la glicina, la dopamina, la acetilcolina y las indolaminas (utilizadas por las células amacrinas).No está claro el transmisor que utilizan las células horizontales, bipolares o interplexiformes. A partir de los fotorreceptores, las señales se transmiten hasta la capa de células ganglionares exclusivamente por conducción electrotónica (potenciales graduados) y no mediante potenciales de acción. La célula ganglionar es la única neurona de la retina capaz de generar un potencial de acción; así se garantiza que las señales de la retina reflejen exactamente la intensidad lumínica, y ello confiere una mayor flexibilidad a las normas de la retina en su respuesta.


Inhibición lateral para potenciar el contraste visual: función de las células horizontales
Las prolongaciones de las células horizontales conectan lateralmente con los cuerpos sinápticos de los fotorreceptores y las dendritas de las células bipolares. Los fotorreceptores situados en el centro de un haz de luz reciben la estimulación máxima, mientras que los periféricos son inactivados por las células horizontales, que, a su vez, se activan por el haz luminoso. Se dice que el entorno se inhibe, mientras que la región central se excita (aunque quizá estos términos no sean precisamente adecuados). Esta es la base de la potenciación del contraste visual. Las células amacrinas también pueden contribuir a potenciar el contraste a través de sus proyección es laterales hacia la capa plexiforme interna. Curiosamente, mientras que algunas células horizontales poseen axones, las células amacrinas no y, en consecuencia, sus propiedades fisiológicas son extraordinariamente complejas.
Algunas células bipolares se excitan con la luz
Algunas células bipolares se despolarizan cuando la luz estimula sus fotorreceptores o receptores asociados, mientras que otras se hiperpolarizan. Esta observación se puede explicar de dos maneras: la primera es que dos células bipolares responden simplemente de modo distinto a la liberación de glutamato por el receptor; la primera se excita con el glutamato y la segunda se inhibe. La segunda explicación es que un tipo de célula bipolar recibiría aferencias (excitadoras) directas del fotorreceptor y el otro aferencias inhibidoras indirectas de una célula horizontal. La excitación de algunas células bipolares y la inhibición de otras podrían, asimismo, contribuir al esquema de inhibición lateral.

Células amacrinas y sus funciones
A través de medios morfológicos e histoquímicos se han identificado unas 30 clases de células amacrinas. Algunas responden enérgicamente al comienzo de la estimulación visual, otras al apagado y otras a la iluminación y al apagado. Existe un tipo celular que solo responde a los estímulos en movimiento. Dada la variedad de neurotransmisores empleados por esta categoría de células, no puede efectuarse ninguna generalización en cuanto a su efecto sobre la neurona efectora.

Células ganglionares y fibras del nervio óptico
La retina dispone de 1,6 millones de células ganglionares; aun así, se cree que existen 100 millones de bastones y 3 millones de conos. Esto significa que sobre cada célula ganglionar de la retina convergen, por término medio, 60 bastones y 2 conos. La población de las células ganglionares se divide en las categorías W, X e Y. Las células ganglionares W representan casi el 40% de la totalidad, son pequeñas y el diámetro del soma es de10 mm; la velocidad de transmisión de los potenciales de acción, 8 m/s, es relativamente lenta. Casi todas sus aferencias provienen de los bastones (a través de las células bipolares y amacrinas) y su campo dendrítico es relativamente amplio. Al parecer, estas células son muy sensibles a los movimientos en el campo visual y, debido a las aferencias predominantes de los bastones, posiblemente se ocupen dela adaptación visual a la oscuridad. Las células ganglionares X son algo más numerosas que las W y representan un 55% de la totalidad. El diámetro del soma varía entre 10 a 15 mm y la velocidad de conducción se aproxima a 14 m/s. Estas células poseen campos dendríticos relativamente pequeños, por lo que representan lugares concretos del campo visual. Cada célula X recibe señales de al menos un cono, por lo que esta clase de célula probablemente se ocupe de la visión en color. Las células ganglionares Y son las más grandes, el diámetro del soma llega hasta 35 mm y conducen con una velocidad aproximada de 50 m/s. Como cabe prever, el campo dendrítico es amplio. No obstante, su número es el más bajo, ya que solo representan un 5% de la totalidad. Estas células responden con rapidez a los cambios en cualquier lugar del campo visual (bien en la intensidad o en el movimiento), pero no especifican con exactitud dónde sucede el cambio.

Excitación de las células ganglionares
Las fibras del nervio óptico están formadas por axones delas células ganglionares. Aun cuando no sean estimuladas, estas células transmiten potenciales de acción con velocidades que oscilan entre 5 y 40 por segundo. Así pues, las señales visuales se superponen a esta descarga de fondo o espontánea. Muchas células ganglionares son particularmente sensibles a los cambios en la intensidad de la luz. Algunas responden con una mayor descarga cuando se acentúa la intensidad lumínica y otras incrementan la descarga al reducirse la intensidad de la luz. Estos efectos obedecen a la presencia de células bipolares despolarizantes e hiperpolarizantes. La capacidad de respuesta a la fluctuación en la intensidad lumínica también está muy desarrollada en las regiones periféricas y foveal de la retina.
Niveles de contraste e inhibición contralateral
Se dice que las células ganglionares responden a los límites del contraste más que al valor absoluto de iluminación. Cuando una luz difusa y plana activa los fotorreceptores, las células bipolares despolarizantes proporcionan una salida excitadora, pero las hiperpolarizantes y las células horizontales generan una salida inhibitoria simultánea. Cuando un estímulo luminoso posee un contraste nítido, en el límite entre luz y oscuridad, se hiperpolariza un fotorreceptor de la zona iluminada y se transmite, a través de esa célula bipolar, una señal despolarizante hasta una célula ganglionar que empieza a descargar con más frecuencia. El fotorreceptor vecino de la región oscura se despolariza y se inactiva la línea célula bipolar-ganglionar. Simultáneamente, una célula horizontal relacionada con el fotorreceptor hiperpolarizado (iluminado) se inactiva porque el fotorreceptor deja de liberar la sustancia transmisora que despolariza la célula horizontal. Así pues, la influencia hiperpolarizante ejercida por esta célula horizontal sobre el fotorreceptor vecino (despolarizado) de la zona oscura desaparece, con lo que el fotorreceptor se despolariza todavía más. La zona oscura se vuelve «más oscura» y la zona de luz se torna «más iluminada» (es decir, se potencia el contraste).
Transmisión de las señales de color por parte de las células ganglionares
Algunas células ganglionares son estimuladas por los tres tipos de conos. Se cree que estas células ganglionares emiten una señal luminosa «blanca». Sin embargo, la mayoría de las células ganglionares son estimuladas por la luz de una longitud de onda e inhibidas por otra. Así, la luz roja puede excitar una determinada célula ganglionar, y la verde, inhibirla; este es el mecanismo oponente del color y se atribuye al proceso empleado para discriminar los colores. Como el sustrato de este proceso está presente en la retina, el reconocimiento y la percepción del color pueden comenzar, de hecho, en la retina en el elemento receptor sensitivo primario.

El ojo: III. Neurofisiología central de la visión

Vías visuales
Los axones de las células ganglionares de la retina forman el nervio óptico. Los axones que se originan en la mitad nasal dela retina se decusa por el quiasma óptico y los de la mitad temporal continúan por la cara lateral del quiasma sin cruzarse. Los axones retinianos siguen detrás del quiasma en forma de tracto óptico y la mayoría termina en el núcleo geniculado lateral dorsal. Desde aquí, los axones delas neuronas geniculadas siguen discurriendo en sentido posterior en forma de radiaciones geniculocalcarinas (ópticas) y acaban en la corteza visual (estriada) primaria. Además, los axones de la retina se propagan a otras regiones del encéfalo como: 1) el núcleo supraquiasmático (control del ritmo circadiano); 2) los núcleos pretectales (reflejos fotomotores pupilares); 3) el colículo superior (control de los movimientos oculares rápidos), y 4) el núcleo geniculado lateral ventral.
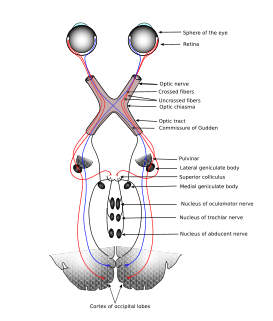
Funciones del núcleo geniculado lateral dorsal (NGLD) del tálamo
Ese núcleo es una estructura laminar que consta de seis capas concéntricas. La más interna es la capa I, y la más superficial, la capa VI. Los axones de la retina que terminan en este núcleo provienen de la retina nasal contralateral y de la retina temporal ipsolateral, y transportan, en consecuencia, una in-formación punto a punto del campo visual contralateral. Las fibras nasales contralaterales terminan en las capas I,IV y VI, y las fibras temporales ipsolaterales, en las capas II, III y V. La información de ambos ojos sigue segregada en el núcleo geniculado lateral dorsal, al igual que las señales provenientes de las células ganglionares X e Y de la retina. La entrada de las células Y termina en las capas I y II, que se denominan capas magno celulares porque contienen neuronas bastante grandes. Esta es una vía de conducción rápida, con ceguera para los colores, pero con una información localizadora eficaz. Las capas III a VI se denominan parvocelulares porque contienen neuronas bastante pequeñas que reciben señales de las células X, que transmiten información del color y de la forma. Por tanto, la información de la retina se procesa a lo largo de al menos dos vías paralelas: 1) una corriente dorsal que trans-porta información de los bastones y las grandes células ganglionares (Y) que dan información específica de la localización y del movimiento, y 2) una corriente ventral que transporta información del color y de la forma desde los conos y las pequeñas células ganglionares (X).

Organización y función de la corteza visual
La corteza visual primaria, o área 17 de Brodmann, también se denomina V-1. Está localizada en la cara medial del hemisferio y reviste las dos paredes del surco calcarino, cerca del polo occipital. Recibe señales visuales de cada ojo y contiene la representación de todo el campo visual contralateral: el campo visual inferior está contenido en el banco superior del surco calcarino, y el superior, en el inferior. La porción macular de la retina se representa posteriormente, cerca del polo occipital, y las señales de la retina más periférica alcanzan los territorios más anteriores. La corteza visual secundaria (denominada V-2 a V-5) rodea el área primaria y se corresponde con las áreas 18 y 19de Brodmann, con la circunvolución temporal media (TM) y con las áreas 7a y 37 de Brodmann.

La corteza visual primaria se estructura en capas
Como todas las demás áreas de la neocorteza, la corteza visual primaria se organiza en seis capas horizontales. Las fibras geniculadas de entrada, de tipo Y, terminan, principalmente, en una subdivisión de la capa IV denominada IVca, mientras que las de tipo X terminan principalmente en las capas IVa y IVcb.
En V-1 también se observa una organización vertical en columnas.
La disposición vertical de las neuronas muestra una anchura aproximada de 50 mm y se extiende por todo el espesor de la corteza, desde la superficie pial hasta la sustancia blanca subcortical. Como las señales talámicas terminan en la capa IV, se dispersan por los circuitos locales, subiendo y bajando por la columna.
Intercaladas entre las columnas se disponen las llamadas manchas de color.
Estos agregados neuronales responden de manera específica a las señales de color mediadas por las columnas corticales circundantes. Las señales visuales de ambos ojos permanecen segregadas a través de las proyecciones desde el núcleo geniculado lateral dorsal hasta V-1. Las células de una columna vertical de la capa IV son las responsables principales de las aferencias de un ojo, y las neuronas de la siguiente columna adyacente responden de preferencia al otro ojo. Estas son las denominadas columnas de dominancia ocular.
Dos vías importantes para el análisis de la información visual
Las relaciones neuronales de la vía de las células Y siguen una corriente más dorsal desde V-1 hasta el área rostral adyacente 18 (V-2) y, después, hasta la corteza parietal. Esta vía señaliza el «lugar de origen» del estímulo y transporta una información localizadora precisa de la imagen visual en el espacio, la forma aproximada de la imagen y su posible movimiento. Por el contrario, la vía más ventral, desde V-1 hasta el áreaV-2 adyacente, y la corteza de la asociación temporal transportan la información de las células X necesaria para el análisis de los detalles visuales. Estas señales se utilizan para reconocerlas texturas, las letras y las palabras, así como el color de los objetos; por tanto, determinan «en qué» consiste el objeto y su significado.
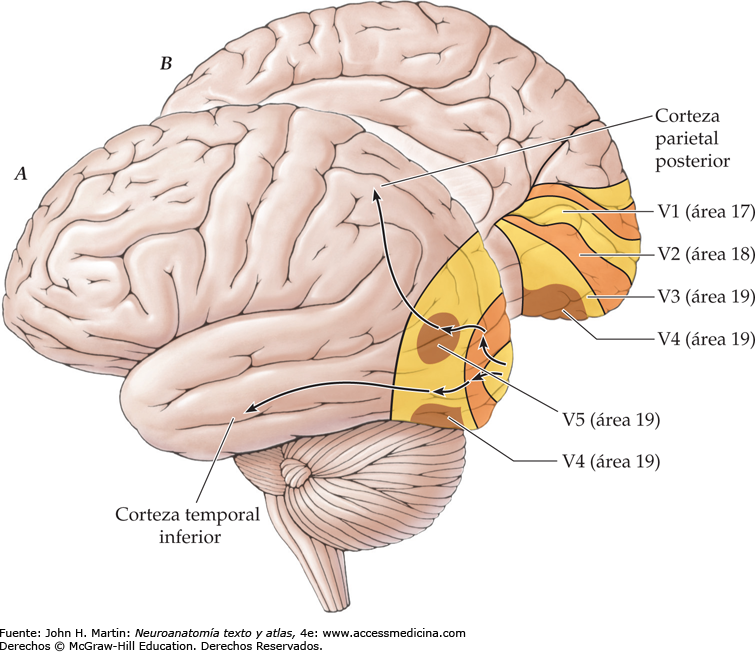
Patrones neuronales de estimulación durante el análisis de una imagen visual
La corteza visual detecta la orientación de las líneas y de los bordes. Anteriormente se comentó (v. capítulo 50) que una delas funciones primordiales del sistema visual es detectar el contraste, principalmente los bordes formados por líneas. Las neuronas de la capa IV de V-1, denominadas células simples, son las principales responsables de que las líneas o los bordes se alineen con una orientación preferida. Otras células de V-1, llamadas células complejas, hacen que las líneas o bordes adopten una orientación preferida, aunque dicha línea pueda desplazarse lateral o verticalmente en una distancia determinada. Una tercera clase de célula, denominada hipercompleja, está localizada, principalmente, en las áreas de asociación visual. Estas células detectan las líneas o bordes con una longitud concreta, una forma angulada específica o un rasgo relativamente complejo. Las neuronas de los diferentes tipos de la corteza visual participan en algunos circuitos organizados en serie y en vías, en las que la información se transmite de forma paralela. Estas dos categorías de organización funcional son importantes para la visión normal.
Detección del color
El color se detecta por medio del contraste. A menudo, el color contrasta con una porción blanca de la escena, que es la base del concepto de constancia del color expuesto en el capítulo50. El contraste del color se detecta mediante un proceso antagónico, en el que ciertos colores excitan determinadas neuronas e inhiben otras. La extirpación de V-1 determina una pérdida de la visión consciente. Las personas pueden seguir reaccionando deforma «reflexiva» a los cambios en la intensidad de la luz, los movimientos en la escena visual y los patrones generales de estimulación lumínica. Esta actividad se debe, fundamentalmente, a los centros visuales subcorticales, como los colículos superiores.

Campos visuales; campimetría
El campo visual –el área vista por un ojo– se divide en una porción nasal (medial) y otra temporal (lateral). La exploración del campo visual de cada ojo por separado se conoce como campimetría. El sujeto se fija en un solo punto del centro del campo visual, mientras un segundo punto pequeño entra y sale de dicho campo. Después, el sujeto debe precisar su localización. En la parte del campo visual ocupada por el disco óptico existe una mancha ciega. Cualquier mancha ciega en otra porción del campo visual se denomina escotoma. En la retinitis pigmentaria degeneran partes de la retina y se deposita, en estas zonas, una cantidad excesiva del pigmento melanina. Este proceso suele iniciarse en la retina periférica y luego se propaga centralmente. Efecto de las lesiones de la vía óptica sobre los campos visuales. La interrupción de las fibras que se decusan en el quiasma óptico determina una pérdida del campo visual de la mitad temporal de cada ojo, denominada hemianopsia bitemporal heterónima. La sección de un tracto óptico hace que se pierda el campo nasal del ojo ipsolateral y el temporal del contralateral, estado que se denomina hemianopsiahomónima contralateral. La lesión de las radiaciones ópticas de un hemisferio produce un defecto similar. Estas dos lesiones se diferencian por la presencia o ausencia de los reflejos fotomotores pupilares. Si los reflejos se preservan, la lesión se encuentra en las radiaciones ópticas; si están abolidos, la lesión debe afectar los tractos ópticos que vehiculan las señales de la retina hasta la región pretectal.
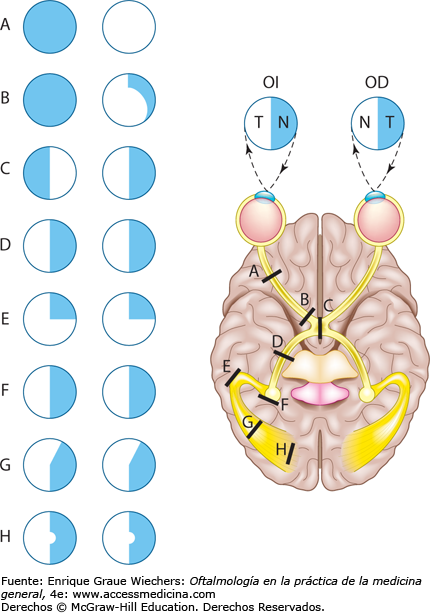
Movimientos oculares y su control
Para que se interprete de manera correcta una escena visual, el cerebro debe desplazar los ojos hasta una posición en que dicha escena se contemple adecuadamente. El movimiento ocular se debe a tres pares de músculos: los rectos medial y lateral, los rectos superior e inferior y los oblicuos superior e inferior. Estos músculos están inervados por motoneuronas delos núcleos de los pares craneales tercero, cuarto y sexto. La actividad de estas motoneuronas depende de diversas áreas del encéfalo, entre ellas las células de los lóbulos frontal, parietal y occipital; la formación reticular del tronco del encéfalo; el colículo superior; el cerebelo; y los núcleos vestibulares. Se consideran tres categorías generales de movimientos oculares: movimientos de fijación, movimientos sacádicos y movimientos de seguimiento.
La fijación consiste en mover los ojos para que una parte concreta del campo visual sea enfocada en la fóvea. La fijación voluntaria está controlada por los campos oculares frontales, el área 8 de Brodmann y un área del lóbulo occipital que representa parte de la corteza visual secundaria (área 19).
Los movimientos sacádicos de los ojos constituyen un mecanismo formado por puntos sucesivos de fijación. Cuando los ojos saltan rápidamente de un objeto a otro, cada salto representa una sacudida. Estos movimientos son rápidos y el cerebro suprime la imagen visual durante el movimiento, por lo que, de ordinario, no se tiene conciencia del movimiento de un punto a otro.

Los movimientos de seguimiento ocurren con la fijación ocular de objetos en movimiento. El sistema regulador de estos movimientos depende de la transmisión de la información visual hasta el cerebelo por diversas vías. Después, el cerebro activa la trayectoria del objeto y las motoneuronas correspondientes desplazan los ojos para que el objeto se mantenga enfocado en la fóvea. Los colículos superiores son los principales responsables de la orientación de los ojos y de la cabeza hacia un estímulo visual (o auditivo). El campo visual se cartografía en el colículo superior de forma independiente al mapa similar creado en la corteza visual. Se piensa que esta actividad está mediada por señales provenientes de las células ganglionares Y de la retina (y quizá también de las células W). El colículo superior también hace que la cabeza y el cuerpo giren hacia el estímulo visual a través de sus proyecciones descendentes por el fascículo tecto espinal. Curiosamente, otras aferencias sensitivas, como la audición o los estímulos somatosensitivos, son canalizadas a través del colículo superior y de sus conexiones descendentes, de forma que el colículo superior cumple una misión integradora global con relación a la orientación delos ojos y del cuerpo hacia diversos estímulos.

Control autónomo de la acomodación y de la apertura pupilar
Las fibras parasimpáticas dirigidas al ojo se originan en el núcleo de Edinger-Westphal y discurren por el nervio motorocular común hasta el ganglio ciliar, donde nacen las fibras posganglionares que llegan al ojo con los nervios ciliares. Las fibras simpáticas se originan en la columna intermedio lateral de la médula y pasan al ganglio cervical superior. Las fibras simpáticas posganglionares viajan con la arteria carótida interna y oftálmica hasta alcanzar finalmente el ojo. Si se modifica el punto de fijación ocular, el poder de enfoque del cristalino se adapta en la dirección adecuada mediante la activación pertinente de la inervación autónoma de los músculos ciliar y esfínter de la pupila de cada ojo. Cuando los ojos cambian el enfoque de un objeto lejano a otro cercano (o viceversa), deben necesariamente converger, para lo que se precisa la activación bilateral de los músculos rectos mediales de cada ojo. Las áreas del encéfalo que regulan los cambios pupilares y la convergencia están suficientemente separadas, puesto que las lesiones alteran una función, pero no la otra. Así, la pupila de Argyll-Robertson es aquella que no muestra reflejos fotomotores normales, pero sí de acomodación. Este tipo de pupila se observa con frecuencia entre las personas aquejadas de sífilis.

