Receptores sensitivos:
Cinco tipos básicos de receptores sensitivos:
- mecanorreceptores, que detectan la compresión mecánica o su estiramiento, o el de los tejidos adyacentes
- termorreceptores, que detectan los cambios en la temperatura, donde algunos de los receptores se encargan del frío y otros del calor
- nocirreceptores (receptores del dolor), que detectan daños físicos o químicos que se producen en los tejidos
- fotorreceptores (electromagnéticos) , que detectan la luz en la retina ocular
- quimiorreceptores, que detectan el gusto en la boca, el olfato en la nariz, la cantidad de oxígeno en la sangre arterial, la osmolalidad de los líquidos corporales, la concentración de dióxido de carbono y otros factores que completen la bioquímica del organismo.
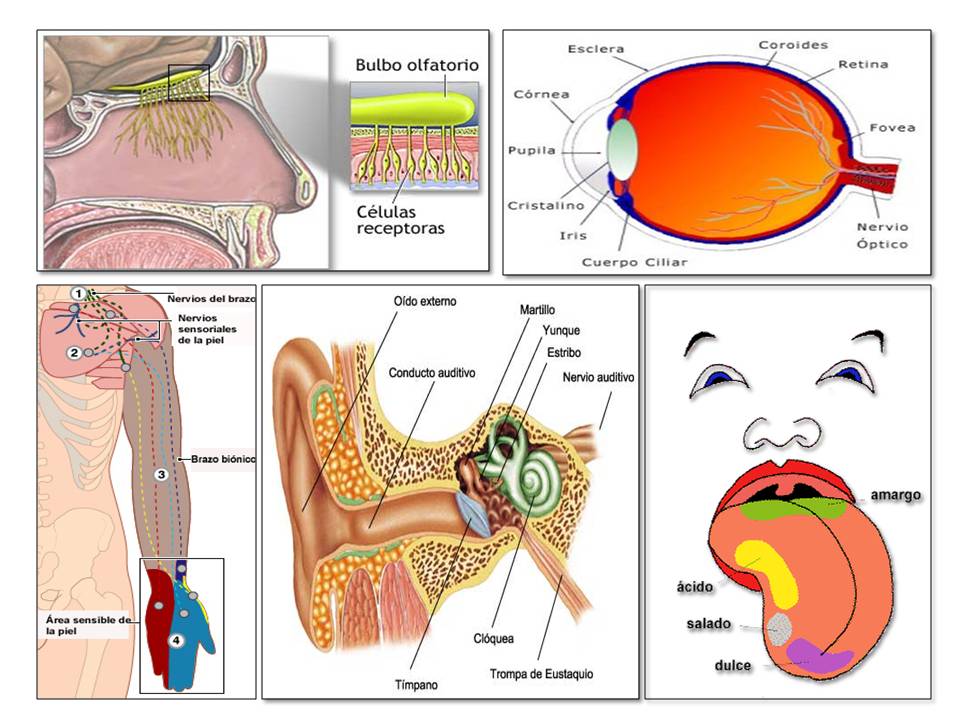
Tipos de receptores sensitivos y estímulos que detectan.

Mecanorreceptores.

Termorreceptores.

Nocirreceptores.

Fotorreceptores.
Quimiorreceptores

Modalidad sensitiva: el principio de la «línea marcada»
Cada uno de los principales tipos sensitivos que podemos experimentar, dolor, tacto, visión, sonido, etc., se llama modalidad de sensación. Con todo, pese al hecho de que nosotros percibimos estas diversas modalidades, las fibras nerviosas únicamente transmiten impulsos. La respuesta señala que cada fascículo nervioso termina en un punto específico del sistema nervioso central y el tipo de sensación vivida cuando se estimula una fibra nerviosa queda determinado por la zona del sistema nervioso a la que conduce esta fibra. Esta especificidad de las fibras nerviosas para transmitir nada más que una modalidad de sensación se llama principio de la línea marcada.
Transducción de estímulos sensitivos en impulsos nerviosos
Corrientes eléctricas locales en las terminaciones nerviosas: potenciales de receptor
Todos los receptores sensitivos tienen un rasgo en común. Cualquiera que sea el tipo de estímulo que les excite, su efecto inmediato consiste en modificar su potencial eléctrico de membrana. Este cambio en el potencial se llama potencial de receptor.
Mecanismos de los potenciales de receptor
Los diversos receptores pueden excitarse siguiendo alguno de los siguientes modos de generar potenciales de receptor: 1) por deformación mecánica del receptor, que estire su membrana y abra los canales iónicos; 2) por la aplicación de un producto químico a la membrana, que también abra los canales iónicos; 3) por un cambio de la temperatura de la membrana, que modifique su permeabilidad, o 4) por los efectos de la radiación electromagnética, como la luz que incide sobre un receptor visual de la retina, al modificar directa o indirectamente las características de la membrana del receptor y permitir el flujo de iones a través de sus canales.
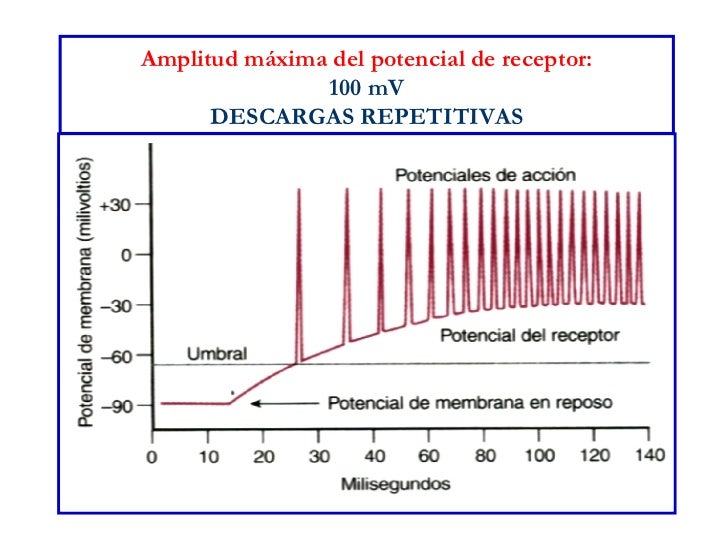
Amplitud del potencial de receptor máximo
La amplitud máxima de la mayoría de los potenciales de receptor sensitivos es de unos 100 mV, pero este valor no se alcanza más que cuando la intensidad del estímulo correspondiente es altísima. Más o menos se trata del mismo voltaje máximo registrado en los potenciales de acción y también es el cambio que sucede cuando la membrana adquiere una permeabilidad máxima a los iones sodio.
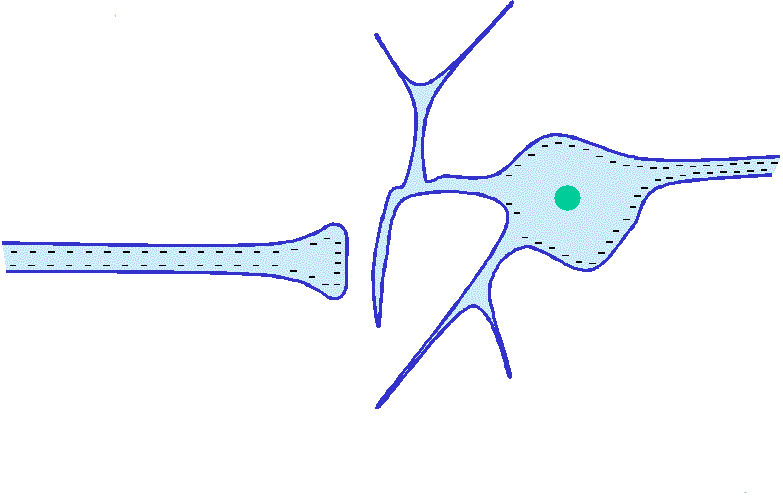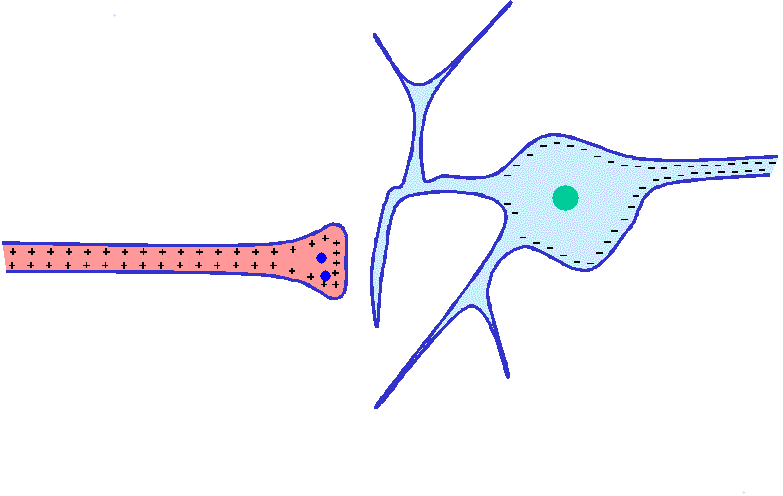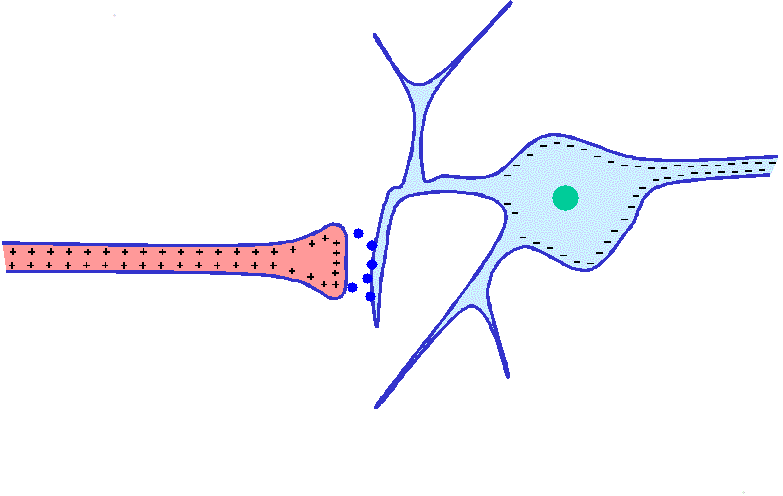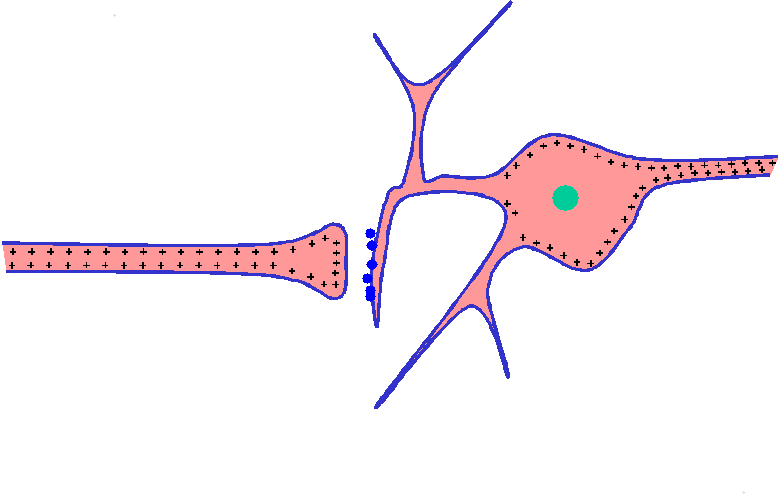
Potencial de receptor del corpúsculo de Pacini: un ejemplo de funcionamiento de un receptor
El corpúsculo de Pacini posee una fibra nerviosa central que recorre su núcleo. Alrededor de esta fibra nerviosa central hay una cápsula compuesta por múltiples capas concéntricas, de manera que la compresión del corpúsculo desde fuera sobre cualquier punto alargará, oprimirá o deformará la fibra central de cualquier otro modo.
La fibra central del corpúsculo de Pacini después de haber retirado todas las capas de la cápsula excepto una. El extremo final con el que acaba en el interior de la cápsula es amielínico, pero la fibra se mieliniza (la cubierta azul que aparece en la figura) poco antes de abandonar el corpúsculo para entrar en un nervio sensitivo periférico.
El mecanismo que produce un potencial de receptor en el corpúsculo de Pacini. Obsérvese la pequeña zona de la fibra terminal que ha quedado deformada por la compresión del corpúsculo, y que los canales iónicos de la membrana se han abierto, lo que permite la difusión de los iones sodio con carga positiva hacia el interior de la fibra. Esta acción crea una mayor positividad dentro de la fibra, que es el «potencial de receptor». A su vez, el citado potencial da lugar a un flujo de corriente formando un circuito local, señalado por las flechas, que se propaga a lo largo de la fibra nerviosa. En el primer nódulo de Ranvier, que aún se halla dentro de la cápsula del corpúsculo de Pacini, este flujo de corriente local despolariza la membrana de la fibra a dicho nivel, lo que a continuación desencadena los potenciales de acción típicos que se transmiten a través de la fibra nerviosa hacia el sistema nervioso central.

Adaptación de los receptores
Todos los receptores sensitivos es su adaptación parcial o total a cualquier estímulo constante después de haber transcurrido un tiempo. Es decir, cuando se aplica un estímulo sensitivo continuo, el receptor responde al principio con una frecuencia de impulsos alta y después baja cada vez más hasta que acaba disminuyendo la frecuencia de los potenciales de acción para pasar a ser muy pocos o muchas veces desaparecer del todo.
Mecanismo de adaptación de los receptores
El mecanismo de adaptación varía con cada tipo de receptor, básicamente lo mismo que la producción de un potencial de receptor constituye una propiedad individual. Por ejemplo, en el ojo, los conos y los bastones se adaptan al modificarse las concentraciones de sus sustancias químicas sensibles a la luz.
En el caso de los mecanorreceptores, el que se ha estudiado con mayor detalle es el corpúsculo de Pacini. En este caso, la adaptación sucede de dos maneras. En primer lugar, el corpúsculo de Pacini es una estructura viscoelástica, por lo que si se aplica de repente una fuerza deformadora sobre uno de sus lados, esta fuerza se transmite al instante directamente al mismo lado de la fibra nerviosa central debido al componente viscoso del corpúsculo, lo que desencadena un potencial de receptor. Sin embargo, en unas pocas centésimas de segundo, el líquido contenido en su interior se redistribuye, de manera que deja de generarse el potencial de receptor. Así pues, este último surge al comienzo de la compresión pero desaparece en cuestión de una pequeña fracción de segundo, aunque siga presente su acción.
El segundo mecanismo de adaptación del corpúsculo de Pacini, mucho más lento en su desarrollo, deriva de un proceso llamado acomodación, que sucede en la propia fibra nerviosa. Esto es, aunque por casualidad la fibra del núcleo central continúe deformada, el extremo de la fibra nerviosa se «acomoda» paulatinamente al estímulo. Esto tal vez obedezca a una «inactivación» progresiva de los canales de sodio en su membrana, lo que significa que el flujo de la corriente de este ion a través suyo hace que se cierren poco a poco, efecto que parece ocurrir en todos o en la mayoría de los canales de sodio de la membrana celular
Los receptores de adaptación lenta detectan la intensidad continua del estímulo: los receptores «tónicos»
Los receptores de adaptación lenta siguen transmitiendo impulsos hacia el cerebro mientras siga presente el estímulo (o al menos durante muchos minutos u horas). Por tanto, mantienen al cerebro constantemente informado sobre la situación del cuerpo y su relación con el medio. Por ejemplo, los impulsos procedentes de los husos musculares y de los aparatos tendinosos de Golgi ponen al sistema nervioso en condiciones de conocer el estado de contracción muscular y la carga soportada por el tendón muscular en cada instante. Otros receptores de adaptación lenta son los siguientes: 1) los pertenecientes a la mácula en el aparato vestibular; 2) los receptores para el dolor; 3) los barorreceptores del árbol arterial, y 4) los quimiorreceptores de los cuerpos carotídeo y aórtico. Debido a su capacidad para seguir transmitiendo información durante muchas horas, o incluso días, también se les denomina receptores tónicos.
Función predictiva de los receptores de velocidad
Si se conoce la velocidad a la que tiene lugar un cambio en la situación corporal, se podrá predecir cuál será el estado del organismo a su juicio unos cuantos segundos o incluso minutos más tarde. Por ejemplo, los receptores existentes en los conductos semicirculares del aparato vestibular del oído detectan la velocidad a la que empieza a girar la cabeza cuando se toma una curva. Con esta información, una persona es capaz de pronosticar el grado de giro durante los 2 s siguientes y corregir el movimiento de las piernas por anticipado para no perder el equilibrio. En este mismo sentido, los receptores situados en las articulaciones o en su proximidad sirven para detectar las velocidades de movimiento que llevan las diferentes partes del cuerpo. Por ejemplo, cuando se corre, la información procedente de los receptores de velocidad articulares le permite al sistema nervioso vaticinar dónde estarán los pies en cualquier fracción exacta del próximo segundo. Por tanto, pueden transmitirse las señales motoras correspondientes a los músculos de las piernas para realizar todas las correcciones de su posición pertinentes como medida preventiva con el fin de no caerse. La pérdida de esta función predictiva impide correr a una persona.
Fibras nerviosas que transmiten diferentes tipos de señales y su clasificación fisiológica
Algunas señales necesitan transmitirse con enorme rapidez hacia el sistema nervioso central o salir de él; si no, la información resultaría inútil. Un ejemplo al respecto lo aportan las señales sensitivas que comunican al cerebro la posición instantánea de las piernas en cada fracción de segundo cuando se corre. En el extremo opuesto, ciertos tipos de información sensitiva, como la que describe un dolor fijo y prolongado, no requieren su envío veloz, por lo que bastará con las fibras de conducción lenta. hay fibras nerviosas de todos los tamaños entre 0,5 y 20 µm de diámetro: cuanto mayor sea este valor, más rápida será su velocidad de conducción. La gama de las velocidades de conducción oscila entre 0,5 y 120 m/s.
Clasificación alternativa empleada por los fisiólogos de la sensibilidad
Ciertas técnicas de registro han permitido dividir las fibras de tipo Aa en dos subgrupos; no obstante, estos mismos métodos no son capaces de distinguir con facilidad entre las fibras Aß y Aγ. Por tanto, los fisiólogos de la sensibilidad emplean a menudo la siguiente clasificación:
Grupo Ia
Fibras procedentes de las terminaciones anuloespirales de los husos musculares (con un diámetro medio de unos 17 µm; son fibras A de tipo a según la clasificación general).

Grupo Ib
Fibras procedentes de los órganos tendinosos de Golgi (con un diámetro medio de unos 16 µm; también son fibras A de tipo a).
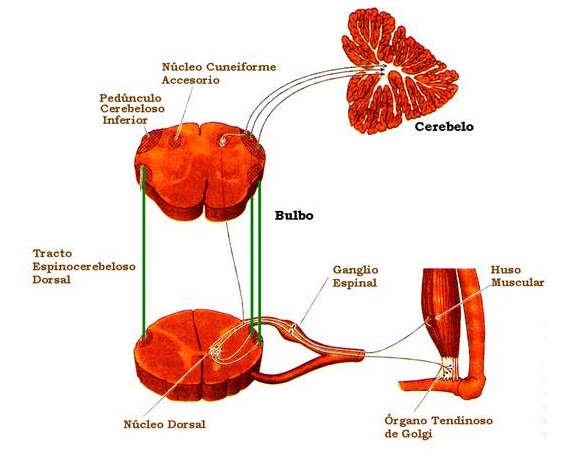
Grupo II
Fibras procedentes de la mayoría de los receptores táctiles cutáneos aislados y de las terminaciones en ramillete de los husos musculares (con un diámetro medio de unos 8 µm; son fibras A de tipo ß y γ según la clasificación general).

Grupo III
Fibras que transportan la temperatura, el tacto grosero y las sensaciones de dolor y escozor (con un diámetro medio de unos 3 µm; son fibras A de tipo d según la clasificación general).

Grupo IV
Fibras amielínicas que transportan las sensaciones de dolor, picor, temperatura y tacto grosero (con un diámetro de 0,5 a 2 µm; son fibras de tipo C según la clasificación general).

Transmisión de señales de diferente intensidad por los fascículos nerviosos: sumación espacial y temporal
Una de las características de toda señal que siempre ha de transportarse es su intensidad: por ejemplo, la intensidad del dolor. Los diversos grados de esta variable pueden transmitirse mediante un número creciente de fibras paralelas o enviando más potenciales de acción a lo largo de una sola fibra. Estos dos mecanismos se llaman, respectivamente, sumación espacial y sumación temporal.
Sumación espacial
El fenómeno de la sumación espacial, por el cual se transmite la intensidad creciente de una señal mediante un número progresivamente mayor de fibras.

Todo el conglomerado formado por las fibras que proceden de una sola con frecuencia cubre una zona de piel cuyo diámetro llega a medir 5 cm. Esta área se llama campo receptor de la fibra. El número de terminaciones es grande en su centro, pero disminuye hacia la periferia. Se puede observar que las fibrillas ramificadas se superponen con las derivadas de otras fibras para el dolor. Por tanto, un pinchazo en la piel suele estimular las terminaciones de muchas diferentes a la vez. Cuando el pinchazo ocurre en el centro del campo receptor de una fibra para el dolor concreta, su grado de estimulación es muy superior a si sucede en la periferia, porque el número de terminaciones nerviosas libres es mucho mayor en dicho caso.
Sumación temporal
Un segundo medio para transmitir señales de intensidad creciente consiste en acelerar la frecuencia de los impulsos nerviosos que recorren cada fibra, lo que se denomina sumación temporal.

Transmisión y procesamiento de las señales en grupos neuronales
El sistema nervioso central está integrado por miles de millones de grupos neuronales; algunos de estos grupos contienen unas cuantas neuronas, mientras que otros presentan una cantidad enorme. Por ejemplo, toda la corteza cerebral podría considerarse un solo gran grupo neuronal. Otros casos similares los ofrecen los diversos componentes de los ganglios basales y los núcleos específicos del tálamo, el cerebelo, el mesencéfalo, la protuberancia y el bulbo raquídeo. Asimismo, toda la sustancia gris dorsal de la médula espinal podría tomarse como un gran grupo de neuronas.

Cada grupo neuronal posee su propia organización especial que le hace procesar las señales de un modo particular y singular, lo que permite que el agregado total de grupos cumpla la multitud de funciones del sistema nervioso.
Transmisión de señales a través de grupos neuronales
Organización de las neuronas para transmitir las señales
Cada fibra que llega se divide cientos o miles de veces, aportando mil fibrillas terminales como mínimo que se esparcen por una gran zona dentro del grupo para hacer sinapsis con las dendritas o los somas de sus neuronas. Estas dendritas también suelen ramificarse y diseminarse a lo largo de cientos o miles de micrómetros en el grupo.
La zona neuronal estimulada por cada fibra nerviosa que entra se llama campo de estimulación. Obsérvese que la neurona más próxima a su «campo» recibe un gran número de terminales derivado de la fibra que entra, pero que esta cantidad es cada vez menor en las neuronas más alejadas.
Estímulos por encima y por debajo del umbral: excitación o facilitación
Un gran número de terminales de llegada ha de actuar sobre la misma neurona a la vez o según una rápida sucesión para provocar esta excitación. Por ejemplo, vamos a suponer que para excitar cualquiera de las neuronas deben descargar seis terminales casi simultáneamente. Cabe observar que la fibra de entrada 1 tiene más de los necesarios para hacer que la neurona a descargue.
Se dice que el estímulo de la fibra de entrada 1 para esta neurona es un estímulo excitador; también se llama estímulo por encima del umbral porque supera el umbral exigido para la excitación. Asimismo, la fibra de entrada 1 aporta terminales a las neuronas b y c, pero no los suficientes para suscitar su excitación. No obstante, la descarga de estos terminales aumenta las posibilidades de que las señales llegadas a través de otras fibras nerviosas de entrada exciten estas neuronas. Por tanto, se dice que los estímulos de estas neuronas están por debajo del umbral, y que las neuronas resultan facilitadas.
En este mismo sentido, para la fibra de entrada 2, el estímulo de la neurona d está por encima del umbral, y los que llegan a las neuronas b y c son estímulos por debajo del umbral, pero facilitadores.

Inhibición de un grupo neuronal
Algunas fibras de entrada inhiben a las neuronas, en vez de excitarlas. Este mecanismo es el opuesto a la facilitación, y el campo de las ramas inhibidoras en su integridad se llama zona inhibidora. El grado de inhibición en el centro de esta área es grande debido al elevado número de terminaciones a dicho nivel y va haciéndose cada vez menor hacia sus bordes.

Divergencia de las señales que atraviesan los grupos neuronales
Muchas veces es importante que las señales débiles que penetran en un grupo neuronal acaben excitando a una cantidad mucho mayor de las fibras nerviosas que lo abandonan. Este fenómeno se llama divergencia. Existen dos tipos fundamentales de divergencia que cumplen unos propósitos totalmente diferentes.
Un tipo de divergencia amplificador esta divergencia significa sencillamente que una señal de entrada se disemina sobre un número creciente de neuronas a medida que atraviesa sucesivos órdenes de células en su camino. Es el tipo de divergencia característico de la vía corticoespinal en su labor de control sobre los músculos esqueléticos, en la que cada célula piramidal grande de la corteza motora es capaz de excitar hasta 10.000 fibras musculares cuando se halla en unas condiciones muy facilitadas. El segundo tipo, es la divergencia en múltiples fascículos. En este caso, la transmisión de la señal desde el grupo sigue dos direcciones. Por ejemplo, la información que llega hasta las columnas dorsales de la médula espinal adopta dos trayectos en la parte baja del encéfalo: 1) hacia el cerebelo, y 2) a través de las regiones inferiores del encéfalo hasta el tálamo y la corteza cerebral. Análogamente, en el tálamo casi toda la información sensitiva se transporta a estructuras talámicas aún más profundas y al mismo tiempo hasta regiones puntuales de la corteza cerebral.
Convergencia de señales
La convergencia significa que un conjunto de señales procedentes de múltiples orígenes se reúnen para excitar una neurona concreta.
La importancia de este tipo de convergencia radica en que las neuronas casi nunca se excitan a partir del potencial de acción de un único terminal de entrada. Sin embargo, los potenciales de acción que convergen sobre la neurona desde muchos terminales proporcionan una sumación espacial suficiente para llevar a la célula hasta el umbral necesario de descarga.
La convergencia también puede surgir con las señales de entrada (excitadoras o inhibidoras) derivadas de múltiples fuentes. Por ejemplo, las interneuronas de la médula espinal reciben señales convergentes desde: 1) fibras nerviosas periféricas que penetran en la médula; 2) fibras propioespinales que pasan de un segmento medular a otro; 3) fibras corticoespinales procedentes de la corteza cerebral, y 4) otras vías largas que descienden desde el encéfalo hasta la médula espinal. A continuación, las señales emitidas por las interneuronas convergen sobre las motoneuronas anteriores para controlar el funcionamiento muscular.
Esta convergencia permite la sumación de información derivada de diversas fuentes y la respuesta resultante reúne el efecto acumulado de todos los diferentes tipos de información. La convergencia es uno de los medios importantes que utiliza el sistema nervioso central para relacionar, sumar y clasificar distintas clases de información.

Circuito neuronal con señales de salida excitadoras e inhibidoras
En ocasiones, una señal de entrada en un grupo neuronal hace que una señal excitadora de salida siga una dirección y a la vez otra señal inhibidora vaya hacia otro lugar. Por ejemplo, al mismo tiempo que una señal excitadora se transmite a lo largo de una serie de neuronas en la médula espinal para provocar el movimiento hacia delante de una pierna, otra señal inhibidora viaja a través de una colección distinta de neuronas para inhibir los músculos de la parte posterior de la pierna a fin de que no se opongan al movimiento hacia delante. Este tipo de circuito es característico en el control de todos los pares de músculos antagonistas y se llama circuito de inhibición recíproca.
Prolongación de una señal por un grupo neuronal: «posdescarga»
Una señal que penetra en un grupo suscita una descarga de salida prolongada, llamada posdescarga, cuya duración va desde unos pocos milisegundos hasta muchos minutos después de que haya acabado la señal de entrada.
Posdescarga sináptica
Cuando las sinapsis excitadoras descargan sobre la superficie de las dendritas o del soma en una neurona, surge en ella un potencial eléctrico postsináptico que dura muchos milisegundos, especialmente cuando interviene alguna de las sustancias transmisoras sinápticas de acción prolongada. Como consecuencia de este mecanismo de «posdescarga» sináptica por sí solo, es posible que una única señal de entrada instantánea dé lugar a la emisión de una señal sostenida (una serie de descargas repetidas) de muchos milisegundos de duración.
Circuito reverberante (oscilatorio) como causa de la prolongación de la señal
Uno de los circuitos más importantes del sistema nervioso es el circuito reverberante u oscilatorio. Está ocasionado por una retroalimentación positiva dentro del circuito neuronal que ejerce una retroalimentación encargada de reexcitar la entrada del mismo circuito. Por consiguiente, una vez estimulado, el circuito puede descargar repetidamente durante mucho tiempo.

Características de la prolongación de la señal en un circuito reverberante
El estímulo de entrada puede durar solo 1 ms más o menos, y sin embargo la salida prolongarse muchos milisegundos o incluso minutos. Al principio de la reverberación la intensidad de la señal de salida suele crecer hasta un valor alto y a continuación disminuye hasta llegar a un punto crítico, en el que súbitamente cesa del todo. La causa de esta repentina interrupción de la reverberación reside en la fatiga de las uniones sinápticas que forman el circuito. Superado cierto nivel decisivo, la fatiga reduce la estimulación de la siguiente neurona en esta cadena hasta dejar de alcanzar el nivel del umbral, por lo que de pronto se desintegra el circuito de retroalimentación.
La duración de la señal total antes de detenerse también puede controlarse por medio de la inhibición o facilitación del circuito a través de las señales procedentes de otras partes del cerebro. Estos patrones de las señales de salida se recogen casi con exactitud en los nervios motores que excitan un músculo implicado en un reflejo flexor tras la estimulación dolorosa del pie.
Emisión de señales continúas desde algunos circuitos neuronales
Algunos circuitos neuronales emiten señales de salida de forma continua, incluso sin señales de entrada excitadoras. Al menos dos mecanismos pueden ocasionar este efecto: 1) la descarga neuronal intrínseca continua, y 2) las señales reverberantes continuas.
Descarga continúa ocasionada por la excitabilidad neuronal intrínseca
Las neuronas, igual que otros tejidos excitables, descargan de forma repetida si el nivel del potencial de membrana excitador sube por encima de un cierto valor umbral. Los potenciales de membrana de muchas neuronas son suficientemente altos incluso en situaciones normales como para hacer que emitan impulsos de forma continua. Este fenómeno sucede sobre todo en gran parte de las neuronas del cerebelo, lo mismo que en la mayoría de las interneuronas de la médula espinal. Las frecuencias de emisión de impulsos en estas células pueden aumentar si reciben señales excitadoras o disminuir si son inhibidoras; estas últimas a menudo tienen la capacidad de rebajar la frecuencia de disparo hasta cero.
Señales continúas emitidas desde circuitos reverberantes como medio para transmitir información
Un circuito reverberante que no alcance un grado de fatiga suficiente como para detener la reverberación es una fuente de impulsos continuos. Además, los impulsos excitadores que penetran en el grupo reverberante pueden incrementar la señal de salida, mientras que la inhibición puede reducir o incluso extinguir la señal.
Inestabilidad y estabilidad de los circuitos neuronales
Casi cualquier parte del cerebro posee conexiones directas o indirectas con cualquier otra parte, lo que crea un serio problema. Si la primera porción excita a la segunda, la segunda a la tercera, la tercera a la cuarta y así sucesivamente hasta que al final la señal reexcite la primera porción, está claro que una señal excitadora que penetre a cualquier nivel del cerebro detonaría un ciclo continuo de reexcitaciones por todas partes. Si este ciclo llegara a producirse, el cerebro quedaría inundado por una nube de señales reverberantes sin control alguno, señales que no estarían portando ninguna información pero que, no obstante, sí estarían ocupando los circuitos cerebrales de manera que sería imposible transmitir cualquier señal realmente informativa. Tal efecto acontece en amplias regiones cerebrales durante las convulsiones epilépticas.mecanismos básicos que funcionan a lo largo de todo el sistema nervioso central: 1) los circuitos inhibidores, y 2) la fatiga de las sinapsis.

Circuitos inhibidores como mecanismo para estabilizar la función del sistema nervioso
Dos tipos de circuitos inhibidores sirven para impedir la difusión excesiva de las señales por extensas regiones del encéfalo: 1) los circuitos de retroalimentación inhibidores que vuelven desde el extremo terminal de una vía hacia las neuronas excitadoras iniciales de esa misma vía (estos circuitos existen prácticamente en todas las vías nerviosas sensitivas e inhiben tanto sus neuronas de entrada como las neuronas intermedias cuando el extremo terminal está demasiado excitado), y 2) ciertos grupos neuronales que ejercen un control inhibidor global sobre regiones generalizadas del cerebro (p. ej., gran parte de los ganglios basales ejercen influencias inhibidoras sobre todo el sistema de control muscular).

Fatiga sináptica como medio para estabilizar el sistema nervioso
La fatiga sináptica significa meramente que la transmisión sináptica se vuelve cada vez más débil cuanto más largo e intenso sea el período de excitación cuanto más breve sea el intervalo entre los reflejos flexores sucesivos, menor será la intensidad de la respuesta refleja posterior.
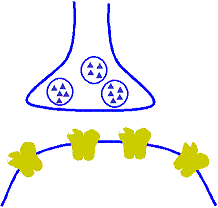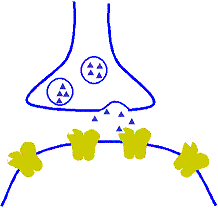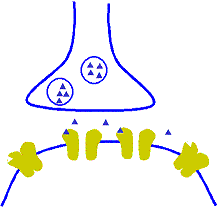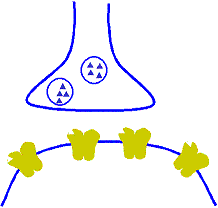
Corrección automática a corto plazo de la sensibilidad de la vía mediante el mecanismo de la fatiga
Cuando están sometidas a un uso excesivo, suelen acabar fatigándose, por lo que desciende su sensibilidad. A la inversa, las que están infrautilizadas se encuentran descansadas y sus sensibilidades aumentan. Por tanto, la fatiga y su recuperación constituyen un medio importante a corto plazo para moderar la sensibilidad de los diferentes circuitos del sistema nervioso. Estas funciones sirven para mantener los circuitos en operación dentro de unos márgenes de sensibilidad que permitan su funcionamiento eficaz.
Cambios a largo plazo en la sensibilidad sináptica ocasionados por la regulación al alza o a la baja de los receptores sinápticos
La sensibilidad a largo plazo de las sinapsis puede cambiar tremendamente si la cantidad de proteínas receptoras presentes en los puntos sinápticos se regula al alza en una situación de baja actividad, y a la baja cuando haya una hiperactividad. El mecanismo de este proceso es el siguiente: las proteínas receptoras están formándose constantemente en el sistema integrado por el retículo endoplásmico y el aparato de Golgi y son introducidas de modo continuo en la membrana sináptica de la neurona receptora. Sin embargo, cuando las sinapsis se emplean demasiado de manera que una cantidad excesiva de sustancia transmisora se combina con las proteínas receptoras, muchos de estos receptores quedan inactivados y retirados de la membrana sináptica.
En efecto, es una suerte que la regulación al alza y a la baja de los receptores, lo mismo que otros mecanismos de control destinados a adecuar la sensibilidad sináptica, corrijan permanentemente esta propiedad en cada circuito hasta el nivel casi exacto necesario para su correcto funcionamiento. Piense por un momento en la seriedad de la situación nada más con que la sensibilidad de unos cuantos de estos circuitos fuera anormalmente alta; entonces cabría esperar la presencia casi continua de calambres musculares, convulsiones, alteraciones psicóticas, alucinaciones, tensión mental u otros trastornos nerviosos. Afortunadamente, los controles automáticos suelen reajustar la sensibilidad de los circuitos de vuelta dentro de unos límites de reactividad controlables en cualquier momento en que empiecen a estar demasiado activos o demasiado deprimidos.
