Diseño general del sistema nervioso
La neurona: unidad funcional básica del sistema nervioso central

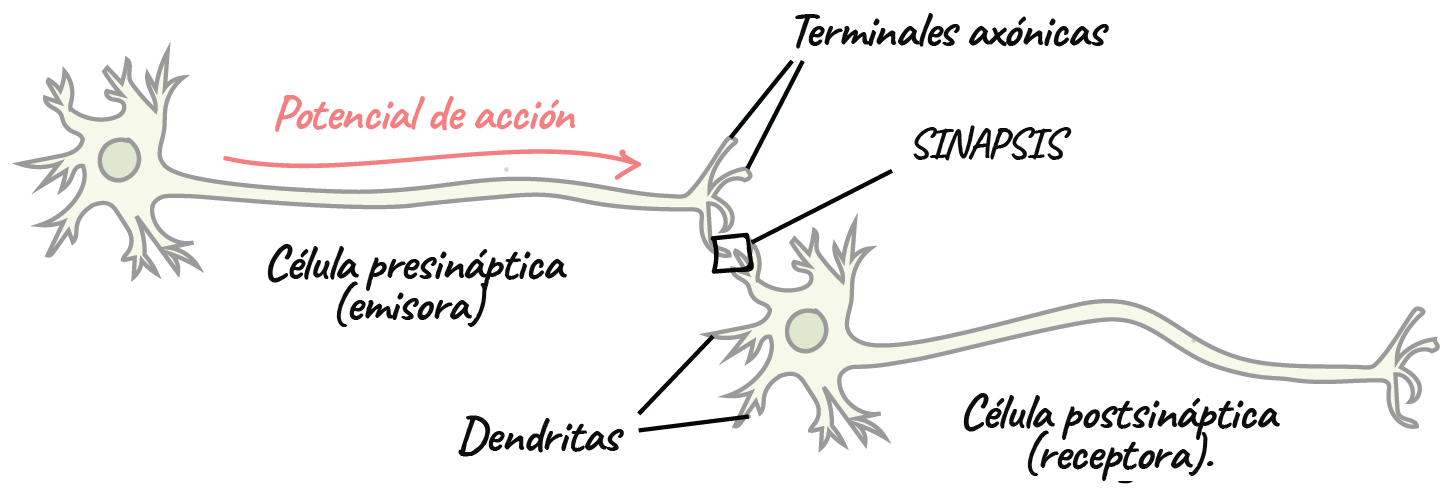
Las señales de entrada llegan a ella a través de las sinapsis situadas fundamentalmente en las dendritas neuronales, pero también en el soma celular. la señal de salida viaja por el único axón que abandona la neurona. A continuación, este axón puede dar origen a numerosas ramas independientes que se dirigen hacia otras zonas del sistema nervioso o de la periferia corporal.
Un rasgo especial de la mayoría de las sinapsis consiste en que normalmente la señal solo circula en sentido anterógrado (desde el axón de una neurona precedente hasta las dendritas en la membrana celular de las neuronas ulteriores).
Porción sensitiva del sistema nervioso: receptores sensitivos
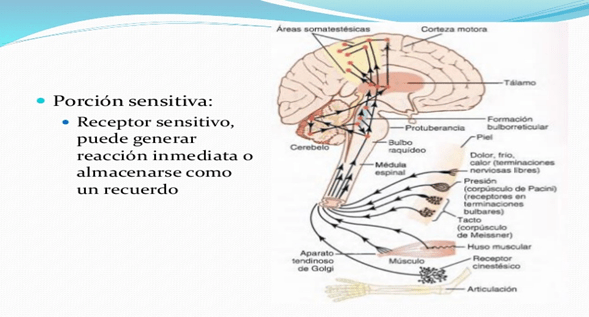
mayoría de las actividades del sistema nervioso se ponen en marcha cuando las experiencias sensitivas excitan los receptores sensitivos, ya sean de carácter visual en los ojos, auditivo en los oídos, táctil en la superficie del organismo o de otros tipos.
Porción motora del sistema nervioso: efectores

Procesamiento de la información: función «integradora» del sistema nervioso
Una de las funciones más importantes del sistema nervioso consiste en elaborar la información que le llega de tal modo que dé lugar a las respuestas motoras y mentales adecuadas. Cuando una información sensitiva importante excita la mente, de inmediato resulta encauzada hacia las regiones motoras e integradoras oportunas del encéfalo para suscitar las respuestas deseadas. Esta canalización y tratamiento de la información se denomina función integradora del sistema nervioso.
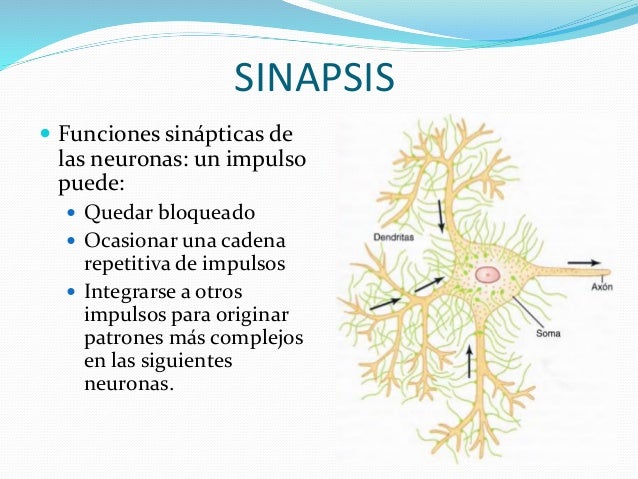
Cometido de las sinapsis en el procesamiento de la información
La sinapsis es el punto de unión de una neurona con la siguiente.
Las sinapsis efectúan una acción selectiva; muchas veces bloquean las señales débiles a la vez que dejan pasar las más potentes, pero en otras circunstancias seleccionan y amplifican ciertas señales débiles y con frecuencia las encarrilan en muchas direcciones en vez de en una sola.
Almacenamiento de la información: memoria
La mayor parte del almacenamiento tiene lugar en la corteza cerebral, pero hasta las regiones basales del encéfalo y la médula espinal pueden conservar pequeñas cantidades de información. La acumulación de la información es el proceso que llamamos memoria, y también constituye una función de las sinapsis.
Principales niveles de función del sistema nervioso central

Sinapsis del sistema nervioso central
La información recorre el sistema nervioso central , por medio de impulsos nerviosos, a través de una sucesión de neuronas, una después de la otra.
Tipos de sinapsis.
La mayoría de las sinapsis utilizadas para la transmisión de señales en el sistema nervioso central del ser humano son sinapsis químicas. En estas sinapsis, la primera neurona segrega un producto químico denominado neurotransmisor que a su vez actúa sobre las proteínas receptoras presentes en la membrana de la neurona siguiente para excitarla, inhibirla o modificar su sensibilidad de algún otro modo.
Entre las mejor conocidas figuran las siguientes: acetilcolina, noradrenalina, adrenalina, histamina, ácido γ-aminobutírico (GABA), glicina, serotonina y glutamato.
Conducción «unidireccional» en las sinapsis químicas
Las sinapsis químicas poseen una característica sumamente importante. Esta característica hace posible que siempre conducen las señales en un solo sentido: es decir, desde la neurona que segrega el neurotransmisor, denominada neurona presináptica, hasta la neurona sobre la que actúa el transmisor, llamada neurona postsináptica. Este fenómeno es el principio de la conducción unidireccional de las sinapsis químicas y se aleja bastante de la conducción a través de las sinapsis eléctricas, que muchas veces transmiten señales en ambos sentidos.
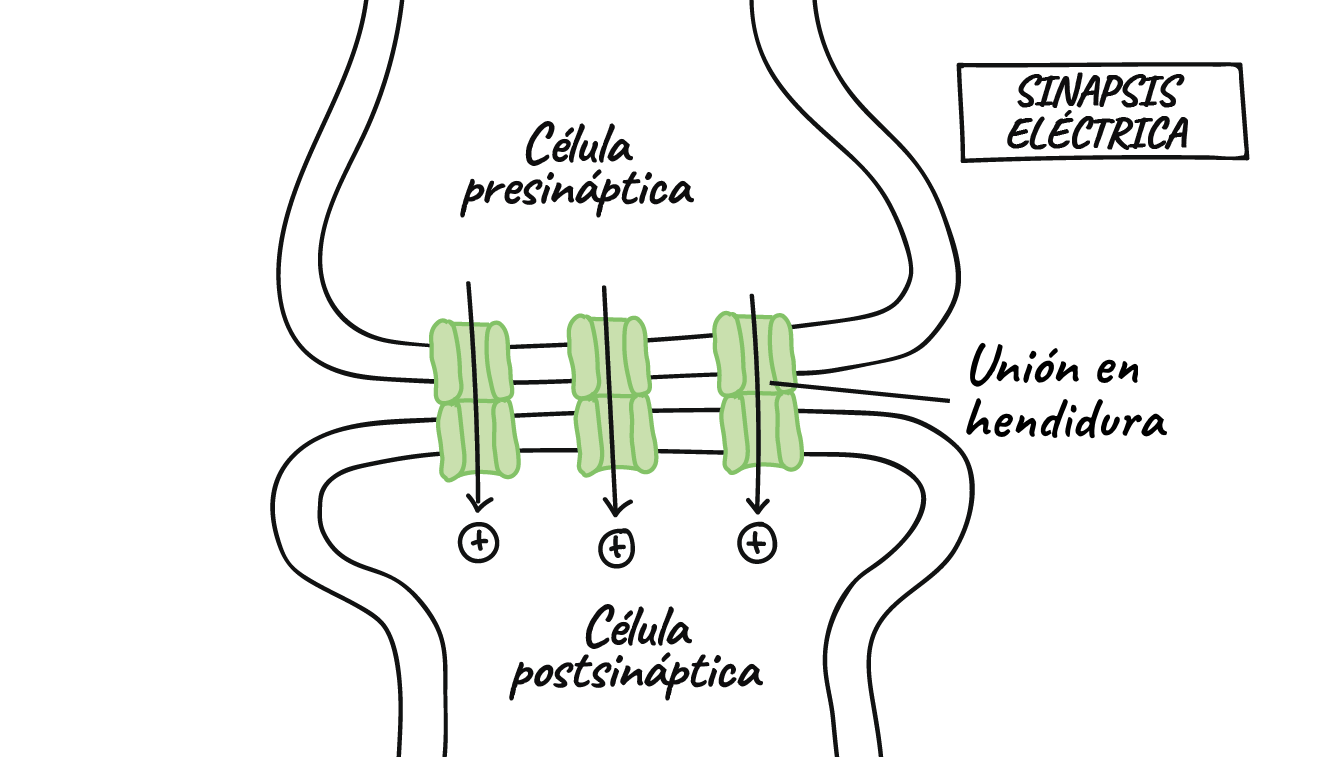
En las sinapsis eléctricas los citoplasmas de las células adyacentes están conectados directamente por grupos de canales de iones llamados uniones en hendidura que permiten el movimiento libre de los iones desde el interior de una célula hasta el interior de la siguiente.
Anatomía fisiológica de la sinapsis.
Está compuesta por tres partes fundamentales: el soma, que es el cuerpo principal de la neurona; el único axón, que se extiende desde el soma hacia un nervio periférico para abandonar la médula espinal, y las dendritas, que constituyen una gran cantidad de prolongaciones ramificadas del soma con unas dimensiones hasta de 1 mm de recorrido hacia las zonas adyacentes en la médula.
Sobre la superficie de las dendritas y del soma de la motoneurona se hallan entre 10.000 y 200.000 diminutos botones sinápticos llamados terminales presinápticos, estando aproximadamente del 80 al 95% en las dendritas y solo del 5 al 20% en el soma.
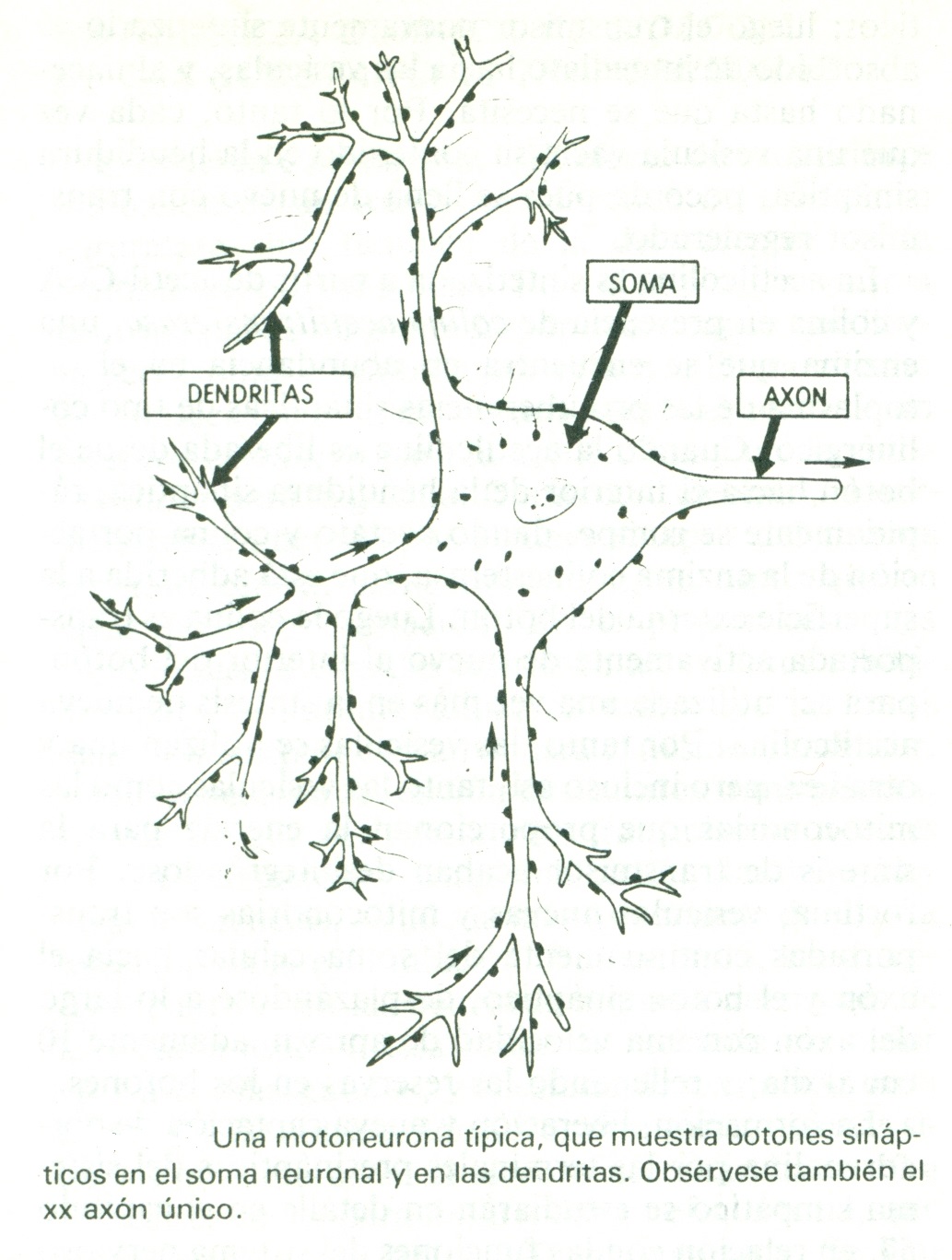
Terminales presinápticos
Se parecen a pequeños botones redondos u ovalados; de ahí que a veces se les llame botones terminales, botones, pies terminales o botones sinápticos. El terminal está separado del soma neuronal postsináptico por una hendidura sináptica cuya anchura suele medir de 200 a 300 angstroms. En él existen dos estructuras internas de importancia para la función excitadora o inhibidora de la sinapsis: las vesículas transmisoras y las mitocondrias. Las vesículas transmisoras contienen el neurotransmisor que, cuando se libera a la hendidura sináptica, excita o inhibe la neurona postsináptica. Excita la neurona postsináptica si la membrana neuronal posee receptores excitadores e inhibe la neurona si contiene receptores inhibidores. Las mitocondrias aportan trifosfato de adenosina (ATP), que a su vez suministra energía para sintetizar más sustancia transmisora.
Mecanismo por el que los potenciales de acción provocan la
liberación del transmisor en los terminales presinápticos:
misión de los iones calcio
La membrana del terminal presináptico se llama membrana presináptica. Contiene una gran abundancia de canales de calcio dependientes de voltaje. Cuando un potencial de acción la despolariza, estos canales se abren y permiten la entrada en el terminal de un número importante de iones calcio. La cantidad de neurotransmisor que sale a continuación hacia la hendidura sináptica desde el terminal es directamente proporcional al total de iones calcio que penetran.

Acción de la sustancia transmisora en la neurona postsináptica: función de las «proteínas receptoras»
La membrana de la neurona postsináptica contiene una gran cantidad de proteínas receptoras Las moléculas de estos receptores poseen dos elementos importantes: 1) un componente de unión que sobresale fuera desde la membrana hacia la hendidura sináptica y donde se fija el neurotransmisor procedente del terminal presináptico, y 2) un componente intracelular que atraviesa toda la membrana postsináptica hasta el interior de la neurona postsináptica. La activación de los receptores controla la apertura de los canales iónicos en la célula postsináptica según una de las dos formas siguientes: 1) por activación de los canales iónicos para permitir el paso de determinados tipos de iones a través de la membrana, o 2) mediante la activación de un «segundo mensajero» que en vez de un canal iónico es una molécula que protruye hacia el citoplasma celular y activa una sustancia o más en el seno de la neurona postsináptica. A su vez, estos segundos mensajeros aumentan o disminuyen determinadas funciones específicas de la célula. Los receptores de neurotransmisores que activan directamente los canales iónicos a menudo se denominan receptores ionotrópicos, mientras que los que actúan a través de sistemas de segundos mensajeros reciben el nombre de receptores metabotrópicos.

Canales iónicos
Los canales catiónicos que transportan iones sodio están revestidos de cargas negativas. Los canales aniónicos, cuando sus diámetros alcanzan las dimensiones suficientes, entran los iones cloruro y los atraviesan hasta el lado opuesto, mientras que los cationes sodio, potasio y calcio quedan retenidos, básicamente porque sus iones hidratados son demasiado grandes para poder pasar.
sistema de segundo mensajero
Los canales iónicos no son idóneos para originar una variación prolongada en las neuronas postsinápticas, porque se cierran en cuestión de milisegundos una vez desaparece la sustancia transmisora. Sin embargo, en muchos casos, se consigue una excitación o una inhibición neuronal postsináptica a largo plazo al activar un sistema químico de «segundo mensajero» en el interior de esta misma célula, y a continuación será este elemento el que genere el efecto duradero.
Existen diversos tipos de sistemas de segundo mensajero. Uno de los más frecuentes recurre a un grupo de proteínas llamadas proteínas G. El complejo de proteínas G inactivas está libre en el citosol y consta de difosfato de guanosina (GDP) más tres elementos: un componente alfa (a), que es la porción activadora de la proteína G, y unos componentes beta (ß) y gamma (γ) que están pegados al componente a. Mientras el complejo de proteínas G está unido a GDP, permanece inactivo. Cuando el receptor es activado por un neurotransmisor, después de un impulso nervioso, experimenta un cambio conformacional, que deja expuesto un sitio de unión para el complejo de proteínas G, que después se une a la porción del receptor que sobresale en el interior de la célula. Este proceso permite que la subunidad a libere GDP y, al mismo tiempo, se una al trifosfato de guanosina (GTP) a la vez que separa de las porciones ß y γ del complejo. El complejo a-GTP desprendido tiene así libertad de movimiento en el citoplasma celular y ejecuta una función o más entre múltiples posibles, según las características específicas de cada tipo de neurona.
Inhibición presináptica
Además de la inhibición originada por las sinapsis inhibidoras que operan en la membrana neuronal, lo que se denomina inhibición postsináptica, muchas veces se produce otro tipo de inhibición en los terminales presinápticos antes de que la señal llegue a alcanzar la sinapsis. Esta clase se llama inhibición presináptica. La inhibición presináptica está ocasionada por la liberación de una sustancia inhibidora en las inmediaciones de las fibrillas nerviosas presinápticas antes de que sus propias terminaciones acaben sobre la neurona postsináptica. En la mayoría de los casos, la sustancia transmisora inhibidora es GABA (ácido γ-aminobutírico).
Funciones especiales de las dendritas para excitar a las neuronas
Campo espacial de excitación de las dendritas amplio
Las dendritas de las motoneuronas anteriores suelen extenderse de 500 a 1.000 μm en todas las direcciones a partir del soma neuronal, y pueden recibir señales procedentes de una gran región espacial en torno a la motoneurona. Esta característica ofrece enormes oportunidades para su sumación desde numerosas fibras nerviosas presinápticas independientes. La mayoría de las dendritas no son capaces de transmitir potenciales de acción, pero sí señales dentro de la misma neurona mediante conducción electrotónica. Relación del estado de excitación de la neurona con la frecuencia de descarga. El «estado excitador» es el nivel acumulado de impulsos excitadores en la neurona. Si en un momento determinado el grado de excitación es más alto que el de inhibición, entonces se dice que existe un estado excitador. A la inversa, si es mayor la inhibición que la excitación, lo que se dice es que hay un estado inhibidor. Cuando el estado excitador de una neurona sube por encima del umbral de excitación, la célula disparará de forma repetida mientras permanezca a ese nivel
Algunas características especiales de la transmisión sináptica
Fatiga de la transmisión sináptica
Cuando las sinapsis excitadoras reciben estímulos repetidos a un ritmo elevado, el número de descargas de la neurona postsináptica es muy alto al principio, pero la frecuencia de disparo va bajando progresivamente en los milisegundos o segundos sucesivos. Este fenómeno se llama fatiga de la transmisión sináptica. La fatiga es una característica importantísima de la función sináptica porque cuando una región del sistema nervioso está hiperexcitada, permite que desaparezca este exceso de excitabilidad pasado un rato.
Efecto de la acidosis o de la alcalosis sobre la transmisión sináptica
La mayoría de las neuronas son muy sensibles a los cambios del pH en los líquidos intersticiales que las rodean. Normalmente, la alcalosis aumenta mucho la excitabilidad neuronal. En cambio, la acidosis disminuye acusadamente la actividad neuronal.
